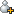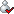图1 细菌细胞从外部环境中去除Cr(Ⅵ)的可能路径
Srinath等发现芽孢杆菌B. circulans,B. megaterium和B.coagulans具有很强的铬络合能力,同时,死细胞在金属去除的过程中不受毒性限制。因此与活细胞相比,死细胞对铬的吸附效果更好,并且死细胞作为生物吸附剂,不需要营养素,吸附的金属离子易于解吸,生物质还可重复利用。实验过程中,通常使用热灭活的细胞进行生物吸附实验,热处理导致细胞壁破裂,释放更多的官能团,从而实现生物质的最大结合能力。生物吸附剂对Cr(Ⅵ)的吸附机理包括在细菌细胞壁中金属与酸性位点之间的离子相互作用和复合物的形成;胞外多糖以表面黏液层的形式结合和积累金属离子,在提高细胞对重金属的吸附能力中发挥了作用。蓝细菌由于比表面积大,黏液体积大,亲和力强和营养要求简单,因此比其他微生物具有更多的吸附优势。同时发现,随着水溶液中Cr(Ⅵ)浓度的增加(测试浓度达20 mg/L),蓝细菌的金属去除率增加。据此推测,由于长期暴露在受重金属污染的地点,这些菌株可能对金属离子产生了耐受性,显示了其生物修复的优势。
1.2细菌类吸附剂对Cr(Ⅲ)的吸附
多项研究表明:使用细菌来源生物吸附剂吸附Cr(Ⅲ)时,细菌合成的胞外多糖不仅为细菌在极端环境环境中提供了保护,而且还提高了其对铬离子的吸附能力。
发现Cr(Ⅲ)的吸附与羰基官能团密切相关。Cr(Ⅲ)浓度的增加会促进该细菌产生胞外多糖,提高对Cr(Ⅲ)的生物去除能力。Cr(Ⅲ)的金属吸附量随着溶液pH、温度和初始金属离子浓度的增加而增加。
2.真 菌
真菌作为生物吸附剂的特征是产生的残留量小、去除速率快,并且能使工业发酵中的真菌废弃生物质增殖。真菌细胞壁成分在生物吸附中起着重要作用,主要包括纤维素、壳聚糖、甲壳素、糖原、葡聚糖和甘露聚糖等。因此,即使没有生理活性,真菌生物质也可以从水溶液中吸附大量重金属。
2.1真菌类吸附剂对Cr(Ⅵ)的吸附
在使用毛霉(Mucor hiemalis)、少根根霉(Rhizopus arrhizus)、黑曲霉(Aspergillus niger)、灰黄青霉(Penicillium griseofulvumMSR1)作为生物吸附剂吸附Cr(Ⅵ)时,均发现Cr(Ⅵ)的去除率随着pH值的降低而增加,随着初始Cr(Ⅵ)浓度上升而增加。动力学研究表明,拟二级模型与实验数据最符合,平衡吸附研究证实Langmuir等温模型对实验数据的拟合效果最好。此外,从ΔH的值可知,生物吸附过程是吸热的。
发现真菌细胞壁中的氨基、羧酸和巯基参与了铬的结合和还原,提出从外部环境中去除Cr(Ⅵ)的可能路径:
1)酸性条件下带正电荷的氨基对带负电的重铬酸根离子的静电吸附;
2)通过形成瞬态的Cr(Ⅵ)硫代酯,硫醇将重铬酸盐还原为Cr(Ⅲ);
3)硫醇氧化为二硫键;
4)Cr(Ⅲ)与氨基配体的螯合作用(图2)。

图2 真菌细胞从外部环境中去除Cr(Ⅵ)的可能路径发现固定化真菌菌丝体比游离真菌菌丝体表现出更高的生物吸附能力和强度。
2.2真菌类吸附剂对Cr(Ⅲ)的吸附
Hanif等利用3种白腐菌(Pleutrotus sajor-cajor、Ganoderma lucidum、Agaricus bitorquis)吸附Cr(Ⅲ),分别进行了分批实验和色谱柱研究,发现生物吸附量随着金属离子浓度(25~200 mg/L)的增加而增加。Langmuir吸附等温线模型和伪二级动力学模型能较好地拟合金属生物吸附数据,硫酸是从死菌生物质中回收金属的最佳洗脱液。Shoaib等研究少根根霉(Rhizopus arrhizus Fisher)对Cr(Ⅲ)的吸附性能时发现,胺基(—NH2)和羟基(—OH)参与了Cr(Ⅲ)与真菌生物质的结合。
3.藻 类
藻类的生物吸附主要归因于金属离子与细胞壁的结合,其细胞壁的主要成分有纤维素、木聚糖、甘露聚糖和海藻酸等,与吸附相关的基团包括羧基、磺酸盐、羟基和氨基等,它们的相对重要性取决于位点的数量以及位点与金属之间的亲和力等因素,主要的金属结合机理包括静电吸引、离子交换和络合作用。
3.1藻类吸附剂对Cr(Ⅵ)的吸附
以藻类作为生物吸附剂吸附Cr(Ⅵ)的机理因使用的藻类不同而存在差异,主要包括静电吸引和吸附—还原2类铬吸附机制。
热力学参数表明,藻类生物对Cr(Ⅵ)的吸附是自发的、吸热的。拟一阶动力学模型能很好地描述动力学数据,速度控制步骤的过程涉及边界层以及粒子内扩散。红外分析表明—COOH、—OH和—NH2基团参与了吸附过程。
Deng等研究刚毛藻(Cladophora albida)对Cr(Ⅵ)的吸附性能时提出其吸附机制符合吸附-还原机制,即:
1)Cr(Ⅵ)阴离子与生物质表面正电荷基团的结合;
2)相邻给电子基团将Cr(Ⅵ)还原为Cr(Ⅲ);
3)正电荷基团与Cr(Ⅲ)离子之间的电子排斥作用使Cr(Ⅲ)离子重新进入水相,或Cr(Ⅲ)与相邻基团形成络合物。
3.2藻类吸附剂对Cr(Ⅲ)的吸附
水中的Cr(Ⅲ)以阳离子形式存在,这些阳离子与藻类中存在的碱金属/碱土金属离子和质子发生离子交换。
Aravindhan等在研究Turbinaria ornate对Cr(Ⅲ)的吸附时发现,其中的羧基与Cr(Ⅲ)络合,提出吸收Cr(Ⅲ)的主要官能团来自多糖。Li等在研究螺旋藻(Spirulina platensis)对Cr(Ⅲ)的吸附特性时提出:在吸附初期,Cr(Ⅲ)被静电吸引到藻细胞表面;然后,通过离子交换与K+、Mn2+、Ca2+、Mg2+、Na+、Fe3+、Zn2+、H+等离子进行可逆或不可逆的化学络合作用,吸附的Cr(Ⅲ)大部分与蛋白质、多糖、脂类等生物配体稳定结合,而只有很小一部分吸附在藻类表面。Onyancha等也认为藻类对Cr(Ⅲ)的吸附是通过离子交换机制进行的,而羧基是主要的结合位点,铬的配位是通过羰基的氧进行的。
4.植 物
植物来源生物吸附剂的优点是成本低、来源多种、原料方便、效率相对较高、对环境的污染很小或几乎没有。其中,与吸附相关的化学成分包括单宁、木质纤维素、多酚和木聚糖等。
4.1植物类吸附剂对Cr(Ⅵ)的吸附
多项研究表明,使用植物来源生物吸附剂吸附Cr(Ⅵ)时,与这些吸附材料结合的铬大部分或全部为Cr(Ⅲ)形式。Dupont等研究了从麸皮中提取的木质纤维素对Cr(Ⅵ)的去除机理,发现吸附反应消耗了大量的质子,使Cr(Ⅵ)还原为Cr(Ⅲ)。木质纤维素部分的氧化与Cr(Ⅵ)还原同时发生,并导致羟基和羧基的形成,并且羧基的形成有助于增加还原铬离子交换位点的数量。而Park等通过XPS光谱发现,含铬生物材料的光谱与Cr(Ⅲ)的光谱完全匹配,由此推测Cr(Ⅵ)还原为Cr(Ⅲ)的反应发生在材料表面。Suksabye等也发现大部分结合在椰壳纤维上的铬都是Cr(Ⅲ)形式,提出椰壳纤维中木质素结构的羰基(—CO)和甲氧基(—O—CH3)基团可能与铬的吸附机理有关。椰壳纤维表面上还原的Cr(Ⅲ)可以通过配位共价键与—CO基团和—O—CH3基团键合,其中甲氧基和羰基氧原子中的1对孤电子可以与Cr(Ⅲ)形成共价键。Vaghetti等在利用巴西松果皮作为生物吸附剂从水溶液中去除Cr(Ⅵ)时提出了多酚对Cr(Ⅵ)的吸附机制:在酸性条件下重铬酸盐将多酚氧化成聚醌,形成的聚醌与重铬酸盐还原生成的Cr(Ⅲ)形成络合物。
4.2植物类吸附剂对Cr(Ⅲ)的吸附
使用植物来源生物吸附剂吸附Cr(Ⅲ)是一个快速的过程,其机理包括静电吸引和络合作用等。
Chojnacka用麦秸秆作为生物吸附剂吸附Cr(Ⅲ),发现生物吸附在10~20 min内达到平衡。并且由于麦秸秆通常含量丰富且成本低,因此可以通过增加吸附剂的浓度来提高废水处理效率。吸附过程的动力学可以用拟二级方程描述,生物吸附平衡可以用Freundlich方程描述,其生物吸附机理为物理吸附。Flogeac等进一步研究了从麦秸中提取的木质纤维素与Cr(Ⅲ)的结合能力,EPR(电子顺磁共振波谱)和XAS(X射线吸收光谱)的微观研究表明,Cr(Ⅲ)在木质纤维素上的吸附是通过形成内表面复合物来实现的。EXAFS(X射线吸收精细结构)研究得出了金属周围精确的局部结构——八面体构型。XANES(X射线吸收近边缘结构)光谱表明,Cr(Ⅲ)与木质纤维素的羧基结合。
5.其 他
除了以上提到的细菌、真菌、藻类和植物来源的天然生物吸附剂以外,为了提高材料对铬离子的吸附性能,大量经天然生物吸附材料改性制成的吸附剂亦被应用。Bingol等先后研究了阳离子表面活性剂修饰的酵母、地衣从水溶液中去除铬酸根阴离子的能力,发现用十六烷基三甲基溴化铵(CTAB)进行改性,可大大改善生物吸附效率。
而植物来源的生物吸附剂中,大多含有木质素,其结构中富含醇羟基和酚羟基,在吸附过程中起着重要作用。也因此,木质素的改性集中在酯化和醚化反应上。据此,Wang等提出通过以下3种方法对木质素进行改性,以提高对铬的吸附效率:
1)将木质素部分解聚或断裂成小片段或芳香大分子单体;
2)通过其他溶剂产生新的化学活性位;
3)酚和醇羟基的化学修饰,以获得木质素基吸附材料的大表面积,强结合位点以及较好的吸附性能。
Yao等发现,枯草芽孢杆菌NX-2衍生的聚谷氨酸(γ-PGA)具有配体功能,通过游离α-羧基与金属离子配位,达到去除Cr(Ⅲ)的效果。除此之外,经过HAc溶解、NaOH沉淀等步骤改性而成的壳聚糖微珠、用壳聚糖与Ustilago(真菌孢子)制成的复合微胶囊、以及利用简单化学处理改性的稻草作为生物吸附剂吸附Cr(Ⅲ),均被证实有良好效果。
03展 望
利用生物吸附法修复铬污染场地一直以来都是研究者不断探索的领域,已有大量文献报道了使用细菌、真菌、藻类和植物来源的生物吸附剂从土壤、水体中去除铬的方法,许多生物质材料在实验室条件下表现出良好的吸附性能,且其低成本、不会造成二次污染以及操作简单的特点,使其在众多重金属修复方法中具有一定优势。但同时生物吸附修复技术仍然存在一些缺点,比如活生物质对铬的耐受能力有限,以其作为生物吸附剂时,仅适用于一些轻度或中度的铬污染场地。除此之外,生物吸附的过程还容易受到外部环境的干扰。
因此,为了充分利用生物吸附剂修复铬污染场地,首先需要阐明微生物代谢的理论机制,以便筛选更多的耐铬微生物菌株,并分析微生物菌株在重金属修复中的安全性和可控性。同时,铬污染生物吸附修复的实验室研究表明,吸附过程受到温度、pH、其他金属离子等各种因素的影响,而在实际场地应用中,吸附过程所面临的环境更为复杂易变。因此需要对污染场地进行长期调查。其次,就目前的研究来说,所涉及的吸附过程和机理仍存在一些不确定性,铬修复过程的机理阐释仍需大量努力和实践。最后,大量文献报道证实改性天然生物质材料是提高吸附性能的有效手段。材料改性通过引入更多的功能性活性基团,且通过增加表面积、化学稳定性和机械强度,来提升吸附剂的吸附能力。同时,改性材料更易于实现循环利用,提升吸附材料的经济性并降低对环境的影响。


 铬污染场地生物吸附修复技术:细菌、…钒污染土壤生物修复研究进展铬污染场地修复技术进展地下水氯代烃污染修复技术研究进展生物炭负载纳米零价铁对污染土壤中铜…环保生物处理技术砷污染土壤修复技术综述15种常见土壤地下水修复技术汇总
铬污染场地生物吸附修复技术:细菌、…钒污染土壤生物修复研究进展铬污染场地修复技术进展地下水氯代烃污染修复技术研究进展生物炭负载纳米零价铁对污染土壤中铜…环保生物处理技术砷污染土壤修复技术综述15种常见土壤地下水修复技术汇总